




A dinâmica populacional de ratos determina seu potencial como problema e influencia a estratégia para seu controle. Em adição a este princípio geral a condução das pesquisas em muitos aspectos do comportamento destes roedores tem um suporte nas táticas de administração de campanhas.
Se em determinada área, a quantidade de alimento não é compatível com a população residente, o excesso migra para novos habitats.
Muitas vezes o aumento da morte de ratos jovens se deve, não a um excedente populacional, mas sim pelo frio, uma vez que os ratos, até dois meses de idade, não tem controle sobre o calor de seu corpo. (Urrego e Urueña, 1986).
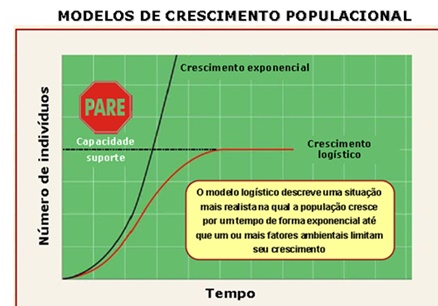
Podemos dizer que as populações têm dois modos básicos de crescimento, crescimento em J (exponencial) e crescimento em S ou sigmóide (logístico).
Estes dois tipos contrastantes podem se combinar ou se modificar dependendo das peculiaridades dos organismos e do meio onde o crescimento é observado.
Modelo de Crescimento Exponencial ou Logarítmico
O modelo de crescimento exponencial descreve uma população que se multiplica por um fator constante (r) durante intervalos de tempo constantes e cujo crescimento depende do número de indivíduos que já existem na população.
O modelo de crescimento exponencial se aplica a populações que crescem sob condições ideais, com recursos ilimitados (sem qualquer ação limitante do ambiente).
Na natureza, o crescimento exponencial é observado apenas durante um curto período de tempo ou em condições especiais.
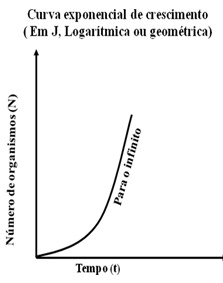
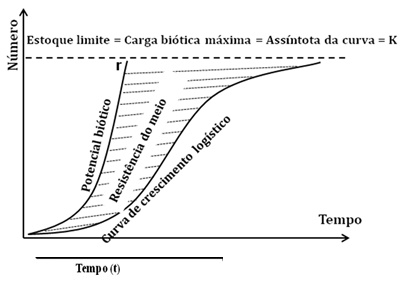
Na forma em J (crescimento exponencial ou geométrico) a densidade aumenta rapidamente, de forma livre onde o índice de crescimento r (o valor máximo de r é o Potencial Biótico é o limite superior de N – crescimento máximo) é constante.
Pode se deter bruscamente quando a densidade poderá permanecer neste nível por algum tempo ou, como ocorre com frequência, se produz uma queda imediata pela ação de um fator ambiental como falta de alimento, espaço ou pela presença de algum predador ou fator climático como gelo, chuva ocasionando morte ou emigração.
Teoricamente o desenvolvimento de uma população que dispõe de recursos alimentares ilimitados e de espaço igualmente ilimitado exprime-se por uma curva exponencial, em J, admitindo-se que o coeficiente de crescimento seja constante.
A curva exponencial corresponde ao potencial biótico. O aumento do número dos indivíduos, se nada houvesse para freá-lo, seria feito segundo uma progressão geométrica. É esta potencia de expansão que corresponde à noção de Potencial Biótico.
Quando uma população entre em um ambiente receptivo novo sua população é baixa e se denomina Início de Sucessão.
A quantidade de vestígios encontrados indica o estágio em que a sucessão se encontra. Poucos vestígios é indicativo de início de sucessão e momento de se usar raticidas pois a população é pequena, os indivíduos não estão muito preocupados com o que vão comer pois a oferta é grande e a competição pequena.

Além disto ocupam grandes nichos, há menor pressão ambiental, a resistência é menor, o stress é menor fazendo com que o sistema hormonal não seja alterado.
Neste momento as populações são mais permanentes não ocorrendo processos migratórios seguidamente a não ser que a população atinja número muito elevado incompatível com a quantidade de alimento oferecida.
Mais tarde a população aumenta, aumentando a competição, o nível de stress, o sistema hormonal é alterado, os nichos diminuem determinando mudanças comportamentais indo em busca de novos ambientes de recepção – emigração.
Com o aumento populacional aumenta o número de indivíduos em reprodução ocupando rapidamente o habitat. São chamados “estrategistas r”, como o rato e o camundongo.
Nesta situação é comum ocorrerem movimentos migratórios com o excedente populacional saindo do habitat de refúgio à procura de novos ambientes reiniciando uma nova sucessão neste novo ambiente – habitat de recepção.
Espécies ditas estrategista r são aquelas que apresentam alta capacidade reprodutiva enquanto que estrategistas k apresentam baixa capacidade reprodutiva.
A ratazana, o rato de telhado e o camundongo são exemplos clássicos de estrategista r: Ninhada de até 13 crias,tempo entre ninhadas 30-50 dias, maturidade sexual com 60 dias, podem entrar em cio a cada 4 ou 5 dias, podem acasalar 1 ou 2 dias depois de parir e gestação de 21 dias. Um único casal pode gerar, em um ano 13.000 descendentes.
Já o camundongo do bosque Apodemus sylvaticus, é exemplo de estrategista k: ninhada de 4-7 filhotes de uma vês ou duas ( no máximo 4) por ano.
Modelo de Crescimento Sigmóide ou Logístico
Como foi discutido anteriormente, em habitats continuamente favoráveis o crescimento logístico de populações de ratos exige contenção regular enquanto populações eruptivas requerem controle intermitente para prevenir erupções. Em ambos os casos, o conhecimento da estrutura social da população é provavelmente para facilitar a administração balanceada.
O sistema social de populações de ratos pode ter um efeito inesperado na tentativa de controle. Em sistemas polígeno, ele é geralmente assumido que a fecundidade da população é limitada pelo número de fêmeas, desde que cada macho possa cobrir muitas fêmeas.
A maioria dos ratos e camundongos é pequena e extremamente fértil. O crescimento logístico requer condições favoráveis contínuas (i.e. alimento, água e esconderijo adequados). Nestas circunstâncias, a população alcançará um nível máximo determinado por fatores dependentes da densidade intraespecífico tais como competição por alimento ou esconderijos.
Um bom exemplo são as condições desfrutadas pelos ratos na palha deixada pelo sistema de plantio direto. Aqui, a manutenção das condições favoráveis são permanentes, seja no momento do plantio, pela disponibilidade de sementes; seja no período de emergência, pela disponibilidade da plântula; seja no período da frutificação, pela disponibilidade direta do produto; seja no momento da colheita, pela disponibilidade dos resíduos ou seja no momento de entre safra, pela disponibilidade da palha. Em todos estes momentos teremos oferta de comida e abrigo.
Onde o crescimento logístico ocorre, medidas de controle (química, mecânica ou ecológica) devem ser mantidos permanentemente, pela constante presença de alimento e esconderijo. Isto porque a existência deste tipo de crescimento indica que as condições são continuamente favoráveis para ratos e deve ser regularmente modificada para tornar o ambiente desfavorável.
Torna-lo desfavorável significa tomarmos medidas que possibilitem a diminuição populacional a níveis aceitáveis, ou seja, de perda significativa de riscos sanitários e econômicos.
Assim, a tomada de decisão é fundamental no manejo do ambiente, aliado ao uso de rodenticidas. Lembrando sempre, que o manejo ambiental só deve ser executado após termos obtidos a redução populacional.
Na forma sigmoide a população aumenta primeiro lentamente (fase de estabelecimento ou crescimento positivo) para depois crescer mais rapidamente (aproximando-se de uma fase exponencial) até um máximo suportado por este ambiente.

Posteriormente vai decrescendo gradualmente à medida em que a resistência ambiental aumenta (fase de aceleração negativa) seja em relação ao alimento, espaço, competição, etc. (fatores dependentes da densidade) até que se alcance um nível mais ou menos equilibrado.
Este nível máximo é chamado de Carga Biótica Máxima (assíntota K).
RESISTÊNCIA AMBIENTAL
Ou seja, em determinado momento em que uma população tem crescimento exponencial pode, em outro, passar a ter um crescimento logístico se equilibrando com o meio.
Gerenciamento de Sinantrópicos se baseia exatamente no conceito de Resistência Ambiental procurando quais fatores ambientais podem ser usados para reduzir o nível populacional da espécie alvo.
Procuramos fazer a população atingir o nível de equilíbrio mais baixo possível e para isto a pressão ambiental tem que ocorrer mais cedo e mais intensamente.
O que observamos é que toda população quando entra em um novo ambiente (habitat de recepção) ela começa a crescer numa exponencial cujo tempo vai depender de fatores dependentes e independentes da densidade.
A partir de um determinado momento dois fatos podem ocorrer:
1. A população declina rapidamente pela falta inesperada e rápida de alimento e esta falta é prolongada – crescimento eruptivo.
2. Fatores ambientais dependentes e independentes da densidade (mudanças climáticas, redução na oferta de alimentos, desratização, etc.) começam a ocorrer e a população começa a sofrer a resistência ambiental entrando num crescimento sigmoide.
Potencial Biótico

Quando o meio não está limitado (isto é, o espaço, alimento, predação, parasitas não exercem um efeito limitativo) o índice de crescimento específico (i.e. o índice de crescimento da população por indivíduo) se faz constante e máximo segundo uma progressão geométrica nas condições microclimáticas existentes e corresponde à noção de potencial biótico de Chapman, 1930 que se exprime por uma curva exponencial.
A capacidade de crescimento é chamado de r e depende da composição de idade em função da capacidade reprodutora – da faixa etária. Assim haverão vários valores de r para cada população segundo a estrutura desta.
O valor máximo de r é o Potencial Biótico e este é freado através da Resistência Ambiental.
O valor de r só é constante em condições controladas pois na natureza as coisas não são bem assim, não é constante pois a fecundidade e mortalidade variam com as condições do meio, a idade dos animais e o alimento e o espaço não existem em quantidades ilimitadas.
Alguns ambientes favorecem espécies a procriar explosivamente. Em ambientes imprevisíveis, tais como árvores frutíferas, o suprimento de fonte pode exceder a demanda, do mesmo modo os sobreviventes de um período rigoroso acha-se em uma região de abundância com melhores condições. Semelhantemente, em um ambiente efêmero os primeiros imigrantes estão livres da escassez.

Então, seu sucesso reprodutivo é desencadeado pela sua habilidade competitiva ou pela densidade populacional. Haverá abundância para ir adiante e o melhor modo de capitalizar sobre isto é produzir um maior número de jovens tanto quanto possível enquanto o ambiente é bom (e para produzir maior número de emigrantes antes que o ambiente se torne ruim).
Este tipo de ambiente é chamado seleção-r, e espécies que seguem este tipo são denominadas seleção-r por evolução (o nome, r, vem da equação logística a qual descreve o potencial de aumento populacional). Roedores tais como ratos e camundongos são estrategistas-r, com taxa reprodutiva explosiva, ou, no mínimo, intermitente, alta densidade populacional mas frequentemente pobre sobrevivência individual.
Em contrate, animais em um ambiente estável ou de sazonalidade previsível utilizam todos os recantos do ambiente. Nestas circunstâncias o suprimento e a demanda serão contrabalanceados e a população será limitada pela disponibilidade de alimento ou outro recurso. A única maneira de ter uma relativa prosperidade em relação aos competidores é ter uma grande área segura de recursos alimentares disponíveis, e isto coloca como um prêmio aos indivíduos de grande habilidade. Sobre estas circunstâncias, a ênfase é na qualidade e não na quantidade.
Os pais terão mais descendentes com segurança se eles inve
tirem pesadamente na proteção de seus filhotes até estarem preparados para entrarem no processo competitivo. Da mesma forma, os jovens estão muito dependentes de seus pais na habilidade competitiva deles, assim, os pais sobre estas circunstâncias deverão investir pesadamente em sua própria capacidade competitiva.
Tais espécies são ditas serem seleção-K ( onde K se refere à capacidade de ocupar o ambiente, também na equação logística). A seleção-K estimula o sucesso individual sobre condições onde os indivíduos estão vivendo em uma população com capacidade de ocupar todo o ambiente.
Em ambos os casos a recompensa – maior sucesso na vida reprodutiva – são a mesma, mas as táticas para a competição são diferentes para diferentes circunstâncias.
Sobre a circunstância de explosão (seleção-r) enfrentadas por lemmingues, p.ex., é crucial produzir filhotes hoje no caso de não haver o amanhã, assim é vantagem procriar bastante antes que os jovens sejam levados à morte prematura diminuindo a possibilidade de vida. Tamanho pequeno pode significar produção em massa, favorecendo muitos jovens não desenvolvidos sobre os menos precoces.
Tamanho pequeno de corpo demanda uma alta taxa metabólica para compensar uma superfície volumétrica desfavorável. Uma alta taxa metabólica resulta em crescimento rápido e reprodução acelerada.
Alguns roedores tem um metabolismo mais rápido do que se poderia prever em relação a seu tamanho, aparentemente uma adaptação a um estilo de vida seleção-r.

O alto metabolismo de fêmea de lemmingues a capacita a ter uma dúzia de filhotes com 42 dias de vida. Para realizar isto, seu metabolismo disputa em relação ao seu tamanho, mas relativamente seleção-K, o camundongo do bosque Apodemus sylvaticus, o qual, com um metabolismo convencional para seu tamanho produz uma ninhada de 4-7 filhotes de uma vês ou duas (no máximo 4) por ano.
A distinção entre r – K é somente uma: uma ratazana é seleção-r em relação à capivara, mas é seleção-K em relação a pequenos roedores do campo. Os ratos urbanos tendem a ser seleção-r. A maioria dos ratos rbanos são pequenos e extremamente férteis.
Muitas espécies se tornam sexualmente maduras aos 2 ou 3 meses de vida e as fêmeas produzem ninhadas de 6 ou 7 jovens após um curto período gestacional de 2 – 3 semanas.
Desta forma, as fêmeas são capazes de serem cobertas após o parto, se tornando prenhas imediatamente após parir, e assim outra ninhada é produzida tão logo a anterior seja desmamada.

Obviamente, esta maximização do processo reprodutivo somente ocorre sobre condições favoráveis, e a longevidade destas condições é que determinarão se ocorrerá um crescimento populacional logístico ou eruptivo.
O crescimento logístico requer condições favoráveis contínuas (i.e. alimento, água e esconderijo adequados). Nestas circunstâncias, a população alcançará um nível máximo determinado por fatores dependentes da densidade intraespecífico tais como competição por alimento ou esconderijos. Um bom exemplo são as condições estáveis desfrutada pelo rato de telhado da Malásia Rattus tiomanicus na plantação de palmáceas no Sudoeste da Asia.
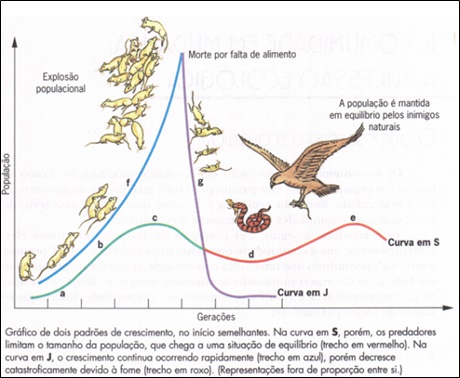
Crescimento eruptivo segue um padrão semelhante ao logístico, com uma partida lenta e acelerando rapidamente entrando numa fase exponencial, mas ao invés de se aproximar de uma assíntota a população de repente cai. Este tipo de crescimento é característico de condições favoráveis instáveis ou descontínuas.
Por exemplo um movimentado ambiente tal como chuvas abaixo da média pode aumentar o período e área sobre a qual aumenta a disponibilidade de comida e abrigo, iniciando com um aumento tanto na duração da estação de produção como na sobrevivência de indivíduos para a próxima estação.
A população de ratos explode e o excedente sai de seu habitat de refúgio e entra em uma área anteriormente desfavorável (habitat de recepção). A queda se inicia quando alimento insuficiente está disponível na estação seguinte para suportar os colonizadores no habitat de recepção e a população rapidamente reverte para um nível que possa ser suportado pelo refúgio. Exemplos de tais crescimentos são as pragas de Microtus arvalis na Europa e Microtus pennsylvanicus nos USA e Mus domesticus na Australia.

Embora estes diferentes tipos de crescimento de populações de roedores surjam do mesmo potencial básico reprodutivo, eles possuem diferentes maneiras de controle.
Onde o crescimento logístico ocorre, medidas de controle (química, mecânica ou ecológica) devem ser mantidos pelo tempo da cultura, mercadorias ou prédios que devem ser protegidos. Isto porque a existência deste tipo de crescimento indica que as condições são continuamente favoráveis para a peste e deve ser regularmente modificada para torna-la desfavorável.
O objetivo é modificar o ambiente de forma que o dano causado seja economicamente insignificante. Um exemplo pode ser a remoção das palmáceas cortadas das plantações durante a colheita, resultando em uma violenta queda nos abrigos do Rattus tiomanicus. Infelizmente neste caso e em muitos outros este tipo de intervenção pré-vazio é economicamente menos viável do que confiar em rodenticidas químicos.
O controle de explosão populacional de ratos requer diferentes estratagemas, visto que os danos ocorrem no tempo de alta população, a manutenção de um controle preventivo deverá ter alta mortalidade a menos que a irupção seja frustrada (p.ex. pela manipulação do habitat).

Assim o controle de focos iruptivos de ratos para prevenir surtos deverá ser com raticidas para cortar o foco no início e para prevenir aumento da população.
A prodigiosa capacidade reprodutiva dos ratos tem consequências além do simples planejamento de campanhas de controle. isto significa também que os predadores não terão sucesso como agentes biológicos controladores da população de ratos. As circunstâncias pelas quais predadores vertebrados podem regular populações de presas são complexas (Sinclair,l989).Entretanto, no contexto de predação como uma medida de controlar ratos, Southern (l979) escreveu a seguinte conclusão geral onde:
(1) os predadores não tem o efeito de frear uma população de presas em expansão, e
(2) seu principal impacto é adiar a recuperação das presas mantendo-a em um nível mais baixo do que poderiam alcançar de outro modo.
As pessoas estão entre os predadores que podem diminuir os ciclos populacionais de ratos, mas forças socioeconômicas estão diminuindo este efeito em algumas comunidades. No Marrocos, Meriones shawi, são roedores reservatórios do protozoário causador da leishmaniose cutânea.
Estes são tradicionalmente controlados por camponeses, mas com a morte de pessoas de comunidades rurais este controle foi relaxado. Consequentemente populações de M. shawi no Marrocos tende a explodir aumentando a prevalência de leishmaniose.
Na Tunisia o principal reservatório de leishmaniose cutânea é o rato do deserto Psammomys obesus, que tem uma adaptação bem acentuada a se alimentar quase que exclusivamente de folhas de plantas da família Chenopodiacea. Estas folhas são baixa em energia e alta em eletrólitos, assim, para maximizar sua ingestão de água os ratos raspam sua epiderme seca antes de ingeri-la.

Anteriormente Chenopodiaceae foram pastejadas pesadamente ao redor dos povoados por cabras e, especialmente, camelos, os quais constituem competidores com os ratos nestas plantas. Com o advento dos veículos o número de camelos se reduziu bastante e o removeu desta competição o que explica a explosão de Psammomys obesus e o aumento de leishmaniose ao redor de novos povoados no deserto.
Irupções de Psammomys também pode ter piorado pela grande destruição de raposas e chacais que os predavam.Controlar Psammomys obesus é especialmente difícil porque, sendo comedor de folhas, ele não come sementes recobertas de veneno. Uma proposta tem sido erradicar sua fonte de alimento ou recolocar competidores como Acacia.
Entretanto, as implicações ecológicas de tais manipulações são desconhecidas e potencialmente imensas.

Mudanças no uso do solo tem tido grande efeito, mas pouco estudada, em populações de ratos. Por exemplo, alternando tratamento de herbicida em fazendas resultou em diferentes movimentos de camundongos da floresta. Além disso as colheitas de cereais resultaram na diminuição de camundongos residentes na área, principalmente através do aumento da predação associada com a perda de abrigo.
A carne moida de camundongos no equipamento de fenação pode explicar como os cavalos ficaram infectados com triquinose, um parasita nematóide que é normalmente transmitido de rato para rato e ocasionalmente é encontrado em seus predadores.
Outra consequencia da alta taxa reprodutiva de ratos sobre condições favoráveis é o aumento de modificações nas gerações e o rápido desenvolvimento de resistência fisiológica aos raticidas anticoagulantes que este gerou. Na verdade, populações de ratos e camundongos na Inglaterra podem também ter desenvolvido uma resistência comportamental a raticidas após décadas de pressão seletiva no rápido crescimento populacional.
A dinâmica populacional de ratos determina seu potencial como problema sanitário e econômico e influencia a estratégia para seu controle. Em adição a este princípio geral a condução das pesquisas em muitos aspectos do comportamento de ratos tem um suporte nas táticas de administração de campanhas.
Teoricamente o desenvolvimento de uma população que dispõe de recursos alimentares ilimitados e de espaço igualmente ilimitado exprime-se por uma curva exponencial, em J, admitindo-se que o coeficiente de crescimento r seja constante, estrategistas-r.
A curva exponencial corresponde ao potencial biótico. O aumento do número dos indivíduos, se nada houvesse para frea-lo, seria feito segundo uma progressão geométrica.
É esta potencia de expansão que corresponde à noção de Potencial Biótico. É preciso por conseguinte opor à taxa de crescimento potencial a taxa de crescimento real de uma população.
É evidente que na natureza as coisas não se passam assim. Primeiramente, o coeficiente r não é geralmente constante, a fecundidade e a mortalidade variam com as condições do meio e a idade dos animais e por fim o alimento e o espaço não existem nunca em quantidade ilimitada.
O matemático francês Verhulst (1845) lançou a hipótese de que o crescimento das populações humanas é representado por uma curva em forma de S, chamada curva logística. Esta curva evidencia a existência de uma densidade máxima de população.
Foi redescoberta por Pearl (1925), que a aplicou ao crescimento de todas as populações animais que dispõem de recursos alimentícios em quantidade fixa mas renovável, é evidente que no caso de recursos alimentícios não renováveis a densidade da população, depois de haver atingido o máximo, declinará e acabará por se anular.
Fatores Dependentes e Independentes da Densidade
Smith (1935) retomando e executando com maior exatidão, particularmente os trabalhos de Howard e Fisk (1911) e os de Thompson (1928), adota uma classificação em fatores independentes e dependentes da densidade.
Os primeiros atuam sobre as populações provocando a destruição de uma percentagem constante de indivíduos, qualquer que seja o número destes. Os segundo destroem uma percentagem de indivíduos que aumenta com a densidade.
Allee (1941) subdivide a categoria dos fatores dependentes da densidade em diretamente dependentes que provocam o aumento da mortalidade quando a densidade aumenta e em fatores inversamente dependentes, que fazem diminuir a mortalidade quando a densidade aumenta.
Visto por este ângulo os fatores independentes são sobretudo os de ordem climática. Uma onda de frio matará uma percentagem da população que não é função de sua densidade. Os fatores dependentes são sobretudo bióticos: competição, predação e o parasitismo exercem efeitos que são função de sua densidade.
FATORES DEPENDENTES
Os chamados fatores dependentes da densidade são aqueles que impedem o crescimento populacional excessivo, devido ao grande número de indivíduos existentes em uma dada população: as disputas por espaço, alimento, parceiro sexual, esconderijo, nidificação, acabam levando à diminuição da taxa reprodutiva e ao aumento da taxa de mortalidade.
O predatismo e o parasitismo são dois outros fatores dependentes da densidade, na medida em que os predadores e parasitas encontram mais facilidade de se espalhar entre os indivíduos de uma população numerosa e o encontro de presas.
Entretanto isto não acontece linearmente, ou seja, em toda e qualquer situação. Existe uma série de outras variáveis que interferem neste conceito que depende da população considerada.
Estes fatores atuam mais fortemente nas populações com baixa vagilidade e valencia ecologica e consequente dificuldade em conseguir alimento e espaço, presença de predadores, dificuldade de esconderijo, isolamento geográfico, etc.
Quando uma população cresce em tamanho, pode esgotar as reservas de alimentos, aumentando a competição entre os membros mais antigos da população e provocando uma alta taxa de mortalidade.
Os Fatores dependentes da densidade podem agir sobre a taxa de crescimento de diferentes maneiras:
A TAXA DE CRESCIMENTO PODE DIMINUIR QUANDO A DENSIDADE AUMENTA.
Estes fatores podem agir sobre a taxa de crescimento de uma população de tres maneiras diferentes:
1) A taxa de crescimento pode diminuir quando a densidade aumenta. Este caso parece muito geral e permite explicar a relativa estabilidade das populações animais. Christian (1961) mostrou que quando o número de ratos aumenta excessivamente nas gaiolas de criação onde o alimento é dado à vontade, os filhotes morrem nas vias genitais da mãe.
A reprodução normal só recomeçará quando o número de ratos tiver voltado a um valor que depende do espaço e não da quantidade de alimento. Strecker e Emlem (1953) descreveram, igualmente no rato, um fenômeno de involução dos órgãos genitais da fêmea em indivíduos que vivem nas gaiolas superpovoadas.
Strecker e Emlem (1953) descreveram, igualmente no rato, um fenômeno de involução dos órgãos genitais da fêmea em indivíduos que vivem nas gaiolas superpovoadas.
Este processo de controle populacional ocorre em diferentes espécies animais. Se colocarmos um único casal de moscas em um frasco com um suprimento fixo de alimentos, haverá um aumento muito grande na população de descendentes, mas apenas no início, pois logo atingirão o limite.
Isso ocorre porque há uma grande competição por alimento entre as larvas, causando uma grande mortalidade em populações muito numerosas. Em populações muito densas, também há uma redução da duração da vida adulta.
Vale lembrar que estas situações acima foram a nível de experimento em área confinada o que não acontece no sistema natural uma vez que o espaço é ilimitado e nos casos de superpopulação o excedente procura outros espaços voluntariamente ou por força da competição.
2) Por vezes a taxa de crescimento permanece aproximadamente constante até a população atingir uma grande densidade, depois diminui bruscamente. Este fato encontra-se nas espécies que apresentam violentas flutuações como os lemingues.
3) Finalmente a taxa de crescimento pode apresentar um máximo para valores médios da densidade. Assim, em algumas aves, como as gaivotas o número de filhotes por casal aumenta com a densidade das aves da colônia, decrescendo depois, logo em seguida a ter passado por um máximo.
A influência da densidade sobre a taxa de crescimento da população pode ter um efeito de retardamento. Após a eliminação de predadores populações crescem rapidamente até a capacidade limite do meio ser alcançada e os animais morrerem de fome, demorando para se recuperar.
Os principais fatores dependentes da densidade são a competição, predação, parasitismo, alimentação e doenças.
FATORES INDEPENDENTES
Muitos autores se recusam a atribuir aos fatores independentes da densidade um papel na regulação das populações. Esta posição parece-nos exagerada porque não leva em conta os resultados obtidos nos estudos auto-ecológicos, resultados que mostram a ação da temperatura, iluminação, etc sobre a duração da vida, fecundidade e muitas outras características dos animais.
O que se pode dizer é que a ação dos fatores climáticos é mais intensa nos pecilotermos que nos homotermos, em função de mecanismos fisiológicos que os tornam relativamente independentes do meio exterior.
Algumas populações tendem a ser autolimitantes porque a taxa de crescimento diminui conforme a densidade aumente. Tais populações tendem a nivelar a densidade antes da saturação e seu crescimento populacional pode ser dito como sendo inversamente dependente da densidade.
Outras não são autolimitantes, mas tendem a crescer em proporção geométrica a menos ou até que sejam freadas por forças exteriores à população (isto é, limitações provenientes de outras populações ou do ecossistema em geral); tais populações podem exaurir suas próprias fontes de energia e as do habitat.
Seu crescimento populacional pode ser dito como sendo independente da densidade, pelo menos até que a densidade se torne muito grande. Quando fracamente reguladas por fatores exteriores à população, estas espécies são sujeitas a grandes oscilações na densidade e podem se tornar problema.
Ainda há um terceiro tipo de relação entre a densidade e a taxa de crescimento. Em algumas espécies a taxa de reprodução é maior em densidades intermediárias do que na densidade baixa ou alta; em outras palavras, tanto a falta como o excesso são limitantes. Este padrão é chamado Tipo de crescimento de Allee.. Ou seja, a taxa de crescimento por indivíduos é primeiro diretamente dependente da densidade e depois inversamente, com um ótimo na densidade intermediária.
CRESCIMENTO ERUPTIVO
Uma variável é o crescimento eruptivo. Esta é a situação mais comumente encontrada, pois os alimentos em dado momento estão em grande quantidade e depois diminuem, são as culturas sazonais, armazéns graneleiros, fábrica de ração, áreas de produção animal, para voltar à abundância mais tarde e assim sucessivamente.
Crescimento eruptivo segue um padrão semelhante ao logístico, com uma partida lenta e acelerando rapidamente entrando numa fase exponencial, mas ao invés de se aproximar de uma assíntota a população de repente cai. Esta queda não significa morte, mas redução populacional por movimentos de emigração, por exemplo. Este tipo de crescimento é característico de condições que variam de favoráveis a desfavoráveis sistematicamente.
Por exemplo, nas épocas de colheita onde os armazéns ficam cheios de alimento por um determinado período. Neste momento há uma imigração para estes locais, aumentando significativamente a população local. Entretanto, esta fartura irá, em determinado momento, diminuir, o que resultará na redução populacional por movimentos emigratórios, saindo de seu habitat de refúgio para novos habitats chamados de habitat de recepção, onde haja alimento suficiente para estes.
Estes movimentos também são percebidos quando, nos habitats de refúgio há um excedente populacional para o alimento existente. O excesso de ratos emigra para novos habitats de recepção.
O controle de explosão populacional de ratos e camundongos requer diferentes estratagemas. Visto que os danos são confinados no tempo, a manutenção de um controle profilático deverá ser permanente, a menos que a irupção seja frustrada (p.ex. pela manipulação do habitat, modificações na forma de armazenamento ou de proteção dos armazéns para se evitar a entrada de ratos).
Assim o controle de focos eruptivos de ratos, deverá estar centrado na previsão e monitoramento de surtos com o uso de raticidas para cortar o foco no início prevenindo a ocorrência de surtos.





